|
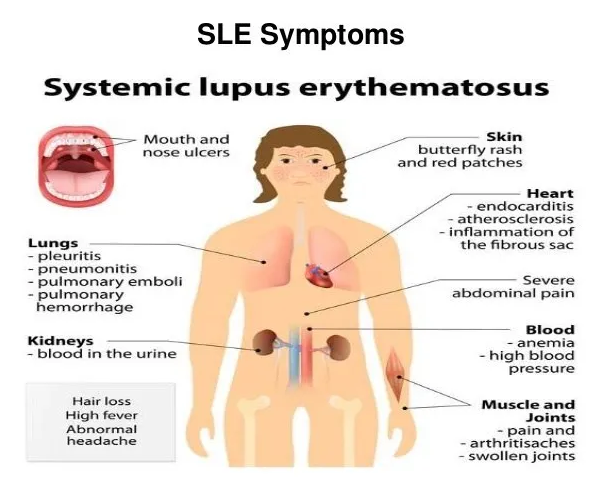
系统性红斑狼疮(SLE)是一种全身性自身免疫性疾病,早期症状最常涉及皮肤和关节,而疾病的发病率和死亡率通常与慢性炎症驱动的心血管事件以及主要器官(尤其是肾脏,神经系统,造血器官)的损害以及免疫抑制剂治疗引起的感染有关。
许多不同的内在和外在因素都会增加SLE患者的感染敏感性。SLE患者的免疫系统功能失调,通常侧重于针对自身,而不是防御入侵的病原体。巨噬细胞,嗜中性粒细胞,T细胞,自然杀伤(NK)和B细胞的缺陷可能对SLE患者感染风险的增加至关重要。
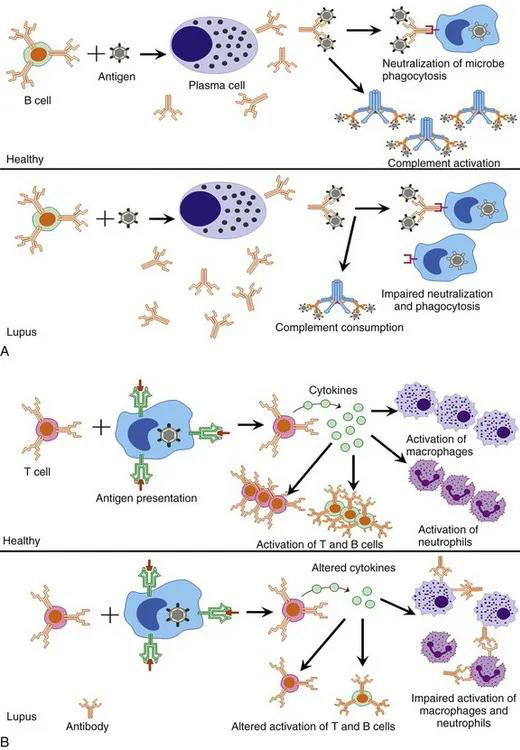 与感染增加相关的系统性红斑狼疮(SLE)的常见受损过程。该图描述了SLE中常见的受损的体液过程(A)和细胞介导的过程(B),它们与感染风险增加有关。 与感染增加相关的系统性红斑狼疮(SLE)的常见受损过程。该图描述了SLE中常见的受损的体液过程(A)和细胞介导的过程(B),它们与感染风险增加有关。
免疫系统
单核/巨噬细胞
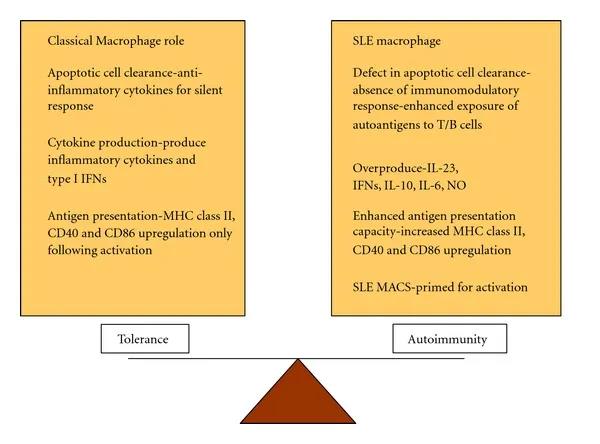
SLE中单核细胞和巨噬细胞均发生表型改变,SLE巨噬细胞显示出凋亡细胞摄取减少,激活状态增强,促炎性和抗炎性巨噬细胞偏斜的改变以及炎性细胞因子如肿瘤坏死因子-α(TNF-α),白介素6(IL-6),白介素10(IL-10)和抗病毒I型干扰素(IFN)。
SLE中巨噬细胞功能失调。患有SLE的患者的免疫系统调节巨噬细胞功能的能力改变。SLE巨噬细胞在凋亡细胞清除方面存在缺陷,过量产生IL-21,IFN,IL-10,IL-6和NO,具有增强的抗原呈递能力并被激活,从而导致自身免疫偏向。
中性粒细胞
中性粒细胞是人类血液中最丰富的白细胞,与SLE有关。SLE中的中性粒细胞功能异常,包括吞噬能力降低,被C1q /钙网蛋白/ CD91介导的凋亡途径清除的能力降低和氧化活性增加。
树突状细胞(DC)
DC为SLE潜在机制的重要参与者,控制单核细胞和DC功能的关键酶之一是血红素加氧酶-1(HO-1)。HO-1在单核细胞和DC中高表达,并且HO-1催化的产物可有助于提高器官移植期间的耐受性。SLE患者或健康对照者的单核细胞中HO-1的表达降低,但DC或CD4+T细胞中的表达却没有降低。
pDC是病毒感染后IFN的主要来源,与SLE的发展相关。SLE患者血清中IFNα水平增加。pDCs为SLE中IFNα产生的重要来源。pDCs释放的IFNα主要发生在疾病早期,来自晚期疾病的pDC在MRL / Mp-Faslpr(lpr)狼疮小鼠模型中无法产生IFNα。
嗜碱性粒细胞
嗜碱性粒细胞是血液中最稀有的免疫细胞群,仅占循环白细胞的1%。临床研究的数据表明,嗜碱性粒细胞有可能被用作SLE疾病活动的生物标记物。源自人类SLE患者的嗜碱性粒细胞能够促进以B细胞产生抗体。 先天性淋巴样细胞(ILC)
ILC代表了先天免疫细胞的新兴家族。最近,在动物模型中发现了ILC和SLE之间的联系,在MRL / Mp-Faslpr(lpr)狼疮小鼠模型中观察到肾浸润性ILC2数量减少,并且随着疾病的进展进一步减少。通过用IL-33进行治疗来恢复ILC2的数量可减少肾小球中免疫细胞的浸润并提高存活率。
补体系统
补体级联的激活由30多种蛋白质组成,可增加趋化性细胞因子的产生,促进抗体的产生,并驱使免疫复合物,凋亡细胞和碎片的清除。早期补体蛋白C1和C4缺乏的个体高度易患SLE(93%的C1q缺乏者和75%的C4缺乏者表现为SLE样症状)。对于补体C1q或C4缺陷小鼠也可导致SLE样疾病。
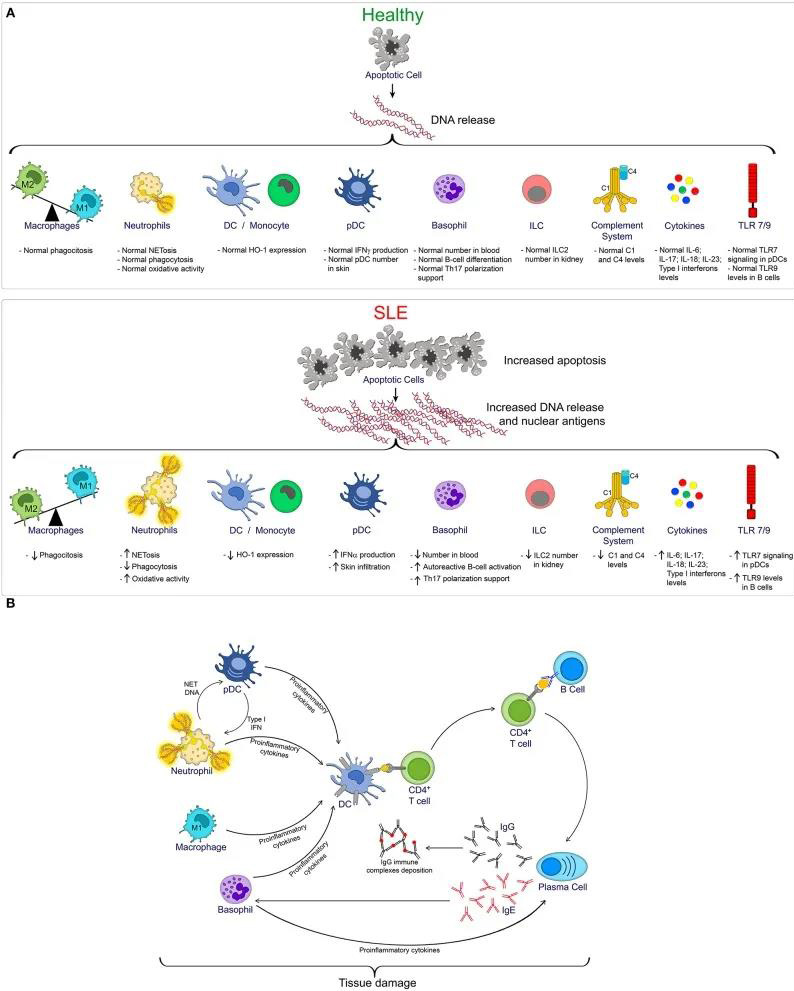
(A) SLE进展过程中受损的先天免疫细胞和途径。在体内平衡状态下(健康),凋亡细胞迅速清除,减少了核抗原的暴露,并降低了自身免疫的风险。然而,如在SLE期间观察到的,凋亡增加以及清除缺陷,增加DNA和核抗原的暴露,促进多种先天免疫细胞的活化。(B) SLE期间先天性和适应性免疫细胞相互作用。
细胞因子
细胞因子主要是由入侵的病原体刺激后,由先天免疫细胞产生的,然后诱导适应性免疫细胞的激活和增殖,以执行关节保护性免疫功能。局部炎症环境会在负反馈中引发抗炎细胞因子的产生,以及调节性T细胞和B细胞的产生,以防止免疫反应过度激活和随后的自身免疫性疾病。但是,SLE患者存在多种细胞因子的数量和功能异常,在疾病发病机理中起着至关重要的作用。这些细胞因子是激活的免疫细胞的下游产物,还可以对免疫细胞的增殖和功能产生不同的影响。它们可以彼此直接或间接地作用,从而构成一个复杂的网络。
 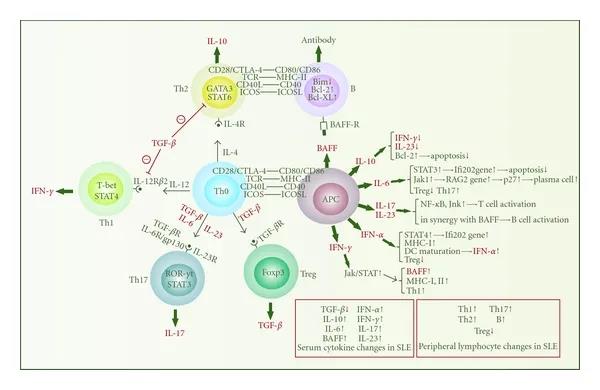
连接固有免疫和适应性免疫的细胞因子网络。该图说明了几种最重要类型的免疫细胞分泌的一些关键细胞因子的功能。APC主要指单核细胞,巨噬细胞和树突状细胞。在SLE中,TGF- β和IL-10主要表现出抗炎作用,而IL-6,BAFF,IFN-α,IFN-γ,IL-17和IL-23则充当促炎细胞因子。
产品推荐
单/多因子检测
| 种属 |
检测技术 |
检测方向 |
货号 |
因子数 |
检测因子 |
| Human |
Luminex |
Cytokine Th1/Th2 |
LX-M5000005L3 |
9 |
GM-CSF, IFN-γ, IL-2, IL-4, IL-5, IL-10, IL-12 (p70), IL-13, TNF-α |
| Human |
Luminex |
Treg cytokines |
LX-171AL003M |
12 |
IL-2,IL-10,IL-12 (p40),IL-12 (p70),IL-19,IL-20,IL-22,IL-26,IL-27 (p28),IL-28A / IFN-λ2,IL-29 / IFN-λ1,IL-35 |
| Human |
Luminex |
Th1/Th2 |
LX-LKTM008 |
11 |
GM-CSF,IFN-gamma,IL-1 beta/IL-1F2,IL-2,IL-4,IL-5,IL-6,IL-10,IL-12 p70,IL-13,TNF-alpha |
| Human |
Luminex |
Th9/Th17/Th22 |
LX-LKTM009 |
17 |
CCL20/MIP-3 alpha,CD40 Ligand/TNFSF5,GM-CSF,IFN-gamma,IL-1 beta/IL-1F2,IL-2,IL-4,IL-5,IL-6,IL-10,IL-12 p70,IL-13,IL-15,IL-17/IL-17A,IL-17E/IL-25,IL-33,TNF-alpha |
| Human |
MSD |
Th17 |
LX-K15085D-1 |
7 |
IL-17A, IL-21, IL-22, IL-23, IL-27, IL-31, MIP-3α |
| Human |
Luminex |
T Cell |
LX-HSTCMAG-28SK-21 |
21 |
Fractalkine/CX3CL1,GM-CSF,IFNγ,IL-1β,IL-2,IL-4,IL-5,IL-6,IL-7,IL-8/CXCL8,IL-10,IL-12(p70),IL-13,IL-17A/CTLA8,IL-21,IL-23,I-TAC/CXCL11,MIP-1α/CCL3,MIP-1β/CCL4,MIP-3α/CCL20,TNFα |
| Human |
Luminex |
Th17 |
LX-HTH17MAG-14K-25 |
25 |
GM-CSF,IFNγ,IL-1β,IL-2,IL-4,IL-5,IL-6,IL-9,IL-10,IL-12(p70),IL-13,IL-15,IL-17A/CTLA8,IL-17E/IL-25,IL-17F,IL-21,IL-22,IL-23,IL-27,IL-28A/IFNλ2,IL-31,IL-33/NF-HEV(mature),MIP-3α/CCL20,TNFα,TNFβ/Lymphotoxin-α(LTA) |
| Human |
Luminex |
CD8+ T Cell |
LX-HCD8MAG-15K-17 |
17 |
sCD137/4-1BB/TNFRSF9,IL-4,IL-5,sFAS/TNFRSF6,sFasL,GM-CSF,Granzyme A ,GranzymeB,IFNγ,IL-2,IL-6,IL-10,IL-13,MIP-1α/CCL3, MIP-1β/CCL4,Perforin TNFα |
| Mouse |
Luminex |
Cytokine Th1/Th2 |
LX-M6000003J7 |
8 |
IL-2, IL-4, IL-5, IL-10, IL-12 (p70), GM-CSF, IFN-γ, TNF-α |
| Mouse |
Luminex |
Cytokine Th17 |
LX-M6000007NY |
6 |
IL-1β, IL-6, IL-10, IL-17A, IFN-γ, TNF-α |
| Mouse |
Luminex |
High Sensitivity T Cell |
LX-MHSTCMAG-70K |
18 |
GM-CSF, IFNγ, IL-1α, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-7, IL-10, IL-12 (p70), IL-13, IL-17A, KC/CXCL1, LIX, MCP-1, MIP-2, TNF-α |
| Mouse |
Luminex |
Th17 |
LX-MTH17MAG-47K |
25 |
CD40 Ligand, GM-CSF, IFN-γ, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-10, IL-12 (p70), IL-13, IL-15, IL-17A, IL-17E/IL-25, IL-17F, IL-21, IL-22, IL-23, IL-27, IL-28B, IL-31, IL-33, MIP-3α/CCL20, TNF-α, TNFβ |
| Mouse |
Luminex |
CD8+ T Cell |
LX-MCD8MAG-48K |
15 |
sCD137, sFas, sFAS ligand, GM-CSF, Granzyme B, IFNγ, IL-2, IL-4, IL-5, IL-6, IL-10, IL-13, MIP-1β, RANTES, TNF-α |
| Mouse |
MSD |
Th17 |
LX-K15246D-1 |
10 |
IL-16,IL-17A,IL-17C,IL-17E/IL-25,IL-17F,IL-21,IL-22,IL-23,IL-31,MIP-3α |
| Rat |
Luminex |
Th17 |
LX-LXSARM-09 |
9 |
GM-CSF、IFN-gamma、IL-1 beta/IL-1F2、IL-2、IL-4、IL-6、IL-10、IL-13、TNF-alpha |
| Rat |
Luminex |
CD8 + T Cell |
LX-LXSARM-08 |
8 |
GM-CSF、IFN-gamma、IL-2、IL-4、IL-6、IL-10、IL-13、TNF-alpha |
| Rat |
Luminex |
Th1/Th2 |
LX-171K1002M |
8 |
IL-2, IL-4, IL-5, IL-10, IL-12 (p70), GM-CSF, IFN-γ, TNF-α |
流式检测
参考文献:
[1]Jennifer C. Byrne, Joan Ní Gabhann, Elisa Lazzari, Rebecca Mahony, Siobhán Smith, Kevin Stacey, Claire Wynne, Caroline A. Jefferies. Genetics of SLE: Functional Relevance for Monocytes/Macrophages in Disease. Jennifer C. Byrne, Joan Ní Gabhann, Elisa Lazzari, Rebecca Mahony, Siobhán Smith, Kevin Stacey, Claire Wynne, Caroline A. Jefferies. Clin Dev Immunol. 2012; 2012: 582352.
[2]Andrés A. Herrada, Noelia Escobedo, Mirentxu Iruretagoyena, Rodrigo A. Valenzuela, Paula I. Burgos, Loreto Cuitino, Carolina Llanos. Innate Immune Cells' Contribution to Systemic Lupus Erythematosus. Front Immunol. 2019; 10: 772.
[3]Ding-Lei Su, Zhi-Min Lu, Min-Ning Shen, Xia Li, Ling-Yun Sun.Roles of Pro- and Anti-Inflammatory Cytokines in the Pathogenesis of SLE. J Biomed Biotechnol. 2012; 2012: 347141.
|












 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




