|
上一期软文中我们对“Immune suppressive landscape in the human esophageal squamous cell carcinoma microenvironment”一文中的图1-4进行了分析,发现在癌症细胞中CD4+、CD8+T细胞,NK细胞与临近组织相比有较明显的变化,且在肿瘤环境中存在免疫抑制。接下来我们进一步针对图5-7进行分析,主要针对巨噬和DC进行了研究。
01 、ESCC中髓细胞的功能组成不同
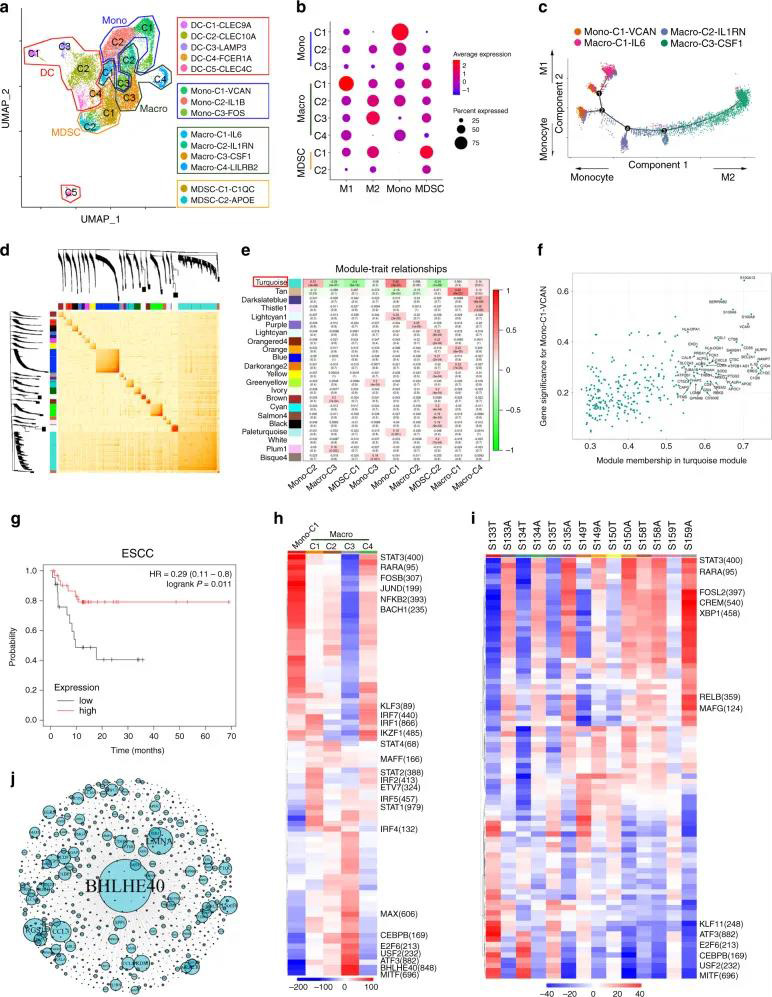
作者进一步进行了髓细胞的无监督聚类。鉴定出14个细胞簇,包括9个单核细胞/巨噬细胞簇和5个DC细胞簇(图5A)。Mono-C1-VCAN显示出很强的单核细胞特征(图5B),为了进一步了解细胞转变,作者使用了Monocle(一种无监督的推理方法)来构建细胞转化的潜在发展轨迹。数据显示单核细胞(Mono-C1-VCAN),M1(Macro-C1-IL6)和M2(Macro-C3-CSF1)位于分支的末端,Macro-C2-IL1RN位于中间(图5C)。接下来,作者使用WGCNA对单核细胞/巨噬细胞进行加权相关网络分析。有趣的是,作者发现Turquoise模块与单核细胞簇Mono-C1-VCAN和Mono-C2-IL1B正相关,而与M2簇Macro-C3-CSF1和MDSC簇MDSC-C1-C1QC,MDSC负相关-C2-APOE(图5D-E)。作者进一步分析了该模块中的基因及其与Mono-C1-VCAN的关联(图5F)以选择最相关的前50个基因,以形成一个集合。这一特征与ESCC存活率的高可能性密切相关(图5G),以及宫颈鳞状细胞癌和肺鳞状细胞癌也出现类似现象,这表明该标记物可作为ESCC和其他组织中鳞状细胞癌的预后生物标志物。
作者进一步应用单细胞调节网络推断和聚类(SCENIC)方法探索可能调节单核细胞M1和M2发育的转录因子。M2中的MITF,BHLHE40,ATF3和USF2被上调,而M1中的IRF转录因子(包括IRF1,IRF7,IRF2,IRF5和PRDM1)被上调。单核细胞簇中的RARA,FOSB和NFKB2大大增加(图5H)。除了S158之外,SCENIC还揭示了大多数肿瘤与相邻组织对之间的分裂(图5I)。转录因子BHLHE40在M2中特异性表达。作者进行了网络分析以鉴定BHLHE40下游基因,并通过Metascape分析了功能(图5J)。BHLHE40可能在TAMs向M2表型诱导中起关键作用,其具体机制有待进一步研究。数据表明,抑制性TAMs在ESCC微环境中的富集可能有助于疾病的进展,阐明了一些与预后相关的令人信服的TF候选基因。
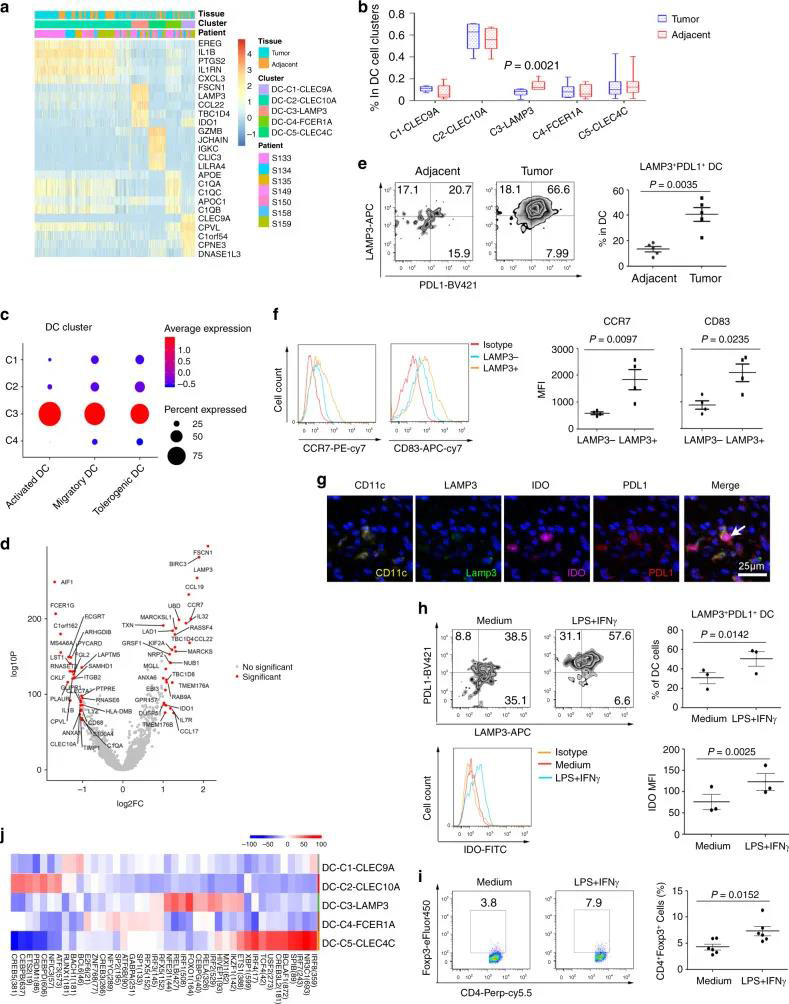
五个DC簇具有高表达水平的CLEC9A,CD1C,FCER1A,LAMP3和CLEC4C,包括常规cDC1(DC-C1-CLEC9A),cDC2(DC-C2-CLEC10A),单核细胞衍生DC(DC-C4-FCER1A) ),LAMP3 + DC(DC-C3-LAMP3)和pDC(DC-C5-CLEC4C)(图6A)。与邻近组织相比,DC-C3-LAMP3富含肿瘤(图6B)。比较了DC的激活和迁移分数,发现LAMP3 + DC与其他DC子集相比具有最高的活性和迁移能力,还发现LAMP3 + DCs丰富了致耐受性的特征(图6C)。LAMP3 + DCs表达了许多调节分子,例如IDO1,EBI3,CD274和IL10(图 6D)。在进行路径富集分析时,发现LAMP3 + DC中上调的基因在细胞因子介导的信号转导,DC细胞分化,白细胞激活,膜运输,抗原加工和呈递的路径中富集(补充图10A),该DC子集支持多功能。
作者通过流式细胞仪验证了LAMP3 + DC。数据显示,与LAMP3-DC相比,LAMP3 + DC的CD83,CCR7和PDL1的表达显着更高(图 6E-F),表明LAMP3 + DCs具有成熟、迁移和调控能力。多色免疫组化染色也证实了肿瘤组织中CD11C + LAMP3 + PDL1 + IDO + DCs的存在(图6G)。有趣的是作者发现IFNγ和LPS刺激诱导DCs表达PDL1和IDO(图6H),并在与CD4 + CD45RA + naïve T细胞共培养时诱导FOXP3表达能力增强(图6I)。提示IFNγ和LPS可诱导树突状细胞产生耐受性。此外,SCENIC分析显示DC亚群可以通过不同的转录因子组来区分(图6J)。LAMP3 + DCs表现出较高的RELB、IRF1、FOXO1和ETS1水平;CEBPD、ETS2、CEBPB、CREB5在cDC2中表达上调。BCL6、BACH1、FLI1和RUNX1在cDC1中高表达,而高水平的SPIB、IRF7和NR3C1与pDCs相关(图6J和补充图10B)。因此,作者进行了网络分析,以通过Metascape鉴定RELB下游基因。数据显示,这些基因与细胞迁移,DC分化和细胞过程的负调控有关(补充图10C-D)。这些结果表明RELB在调节tDC发展中具有重要作用。
02 、ESCC中免疫细胞间的相互作用
细胞间的通信主要通过配体-受体的相互作用,在确定TME和对治疗反应中起关键作用。因此,基于在任何两种类型的肿瘤浸润免疫细胞中的已知配体-受体对的共表达,对潜在的细胞间相互作用进行了系统分析,并在肿瘤与邻近组织之间进行了比较。由于巨噬细胞和Treg在肿瘤中的高浸润水平和重要的免疫调节作用,作者应用了scTHI(https://github.com/miccec/scTHI),这是另一种广泛用于分析巨噬细胞和Tregs相互作用的方法,发现他们中有可比的结果。
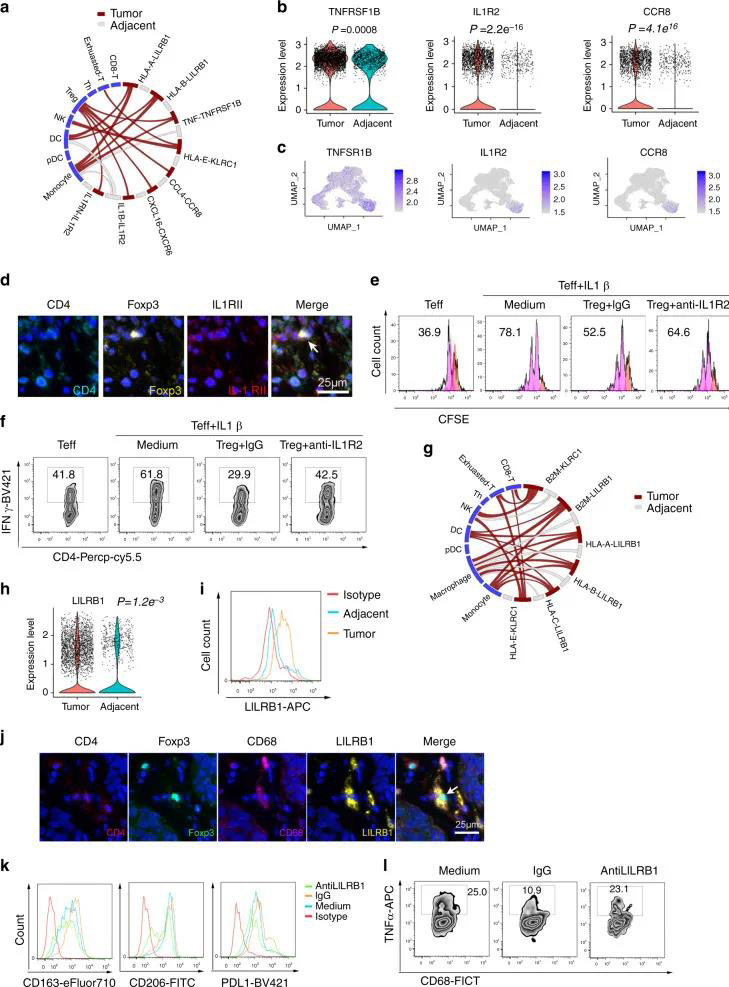
结果发现巨噬细胞和Treg之间的TNF-TNFSF1B,CCL4-CCR8和IL-1β-IL1R2相互作用具有较高的相互作用评分,并且Treg在肿瘤中表达了高水平的TNFSF1B,CCR8和IL1R2(图7A-C)。首先,通过流式细胞术分析了食管鳞癌和邻近组织的IL1R2表达,发现从肿瘤分离的Tregs中IL1R2表达高于邻近组织(补充图11A)。然后,多色IHC还验证了Tregs中的IL1R2表达(图7D)。第三,体外共培养和抗体阻断试验表明,IL1R2对于Treg抑制抑制效应细胞T的IL-1β依赖性激活(例如增殖和IFNγ表达)是必需的(图7E-F,补充图11B-C)。
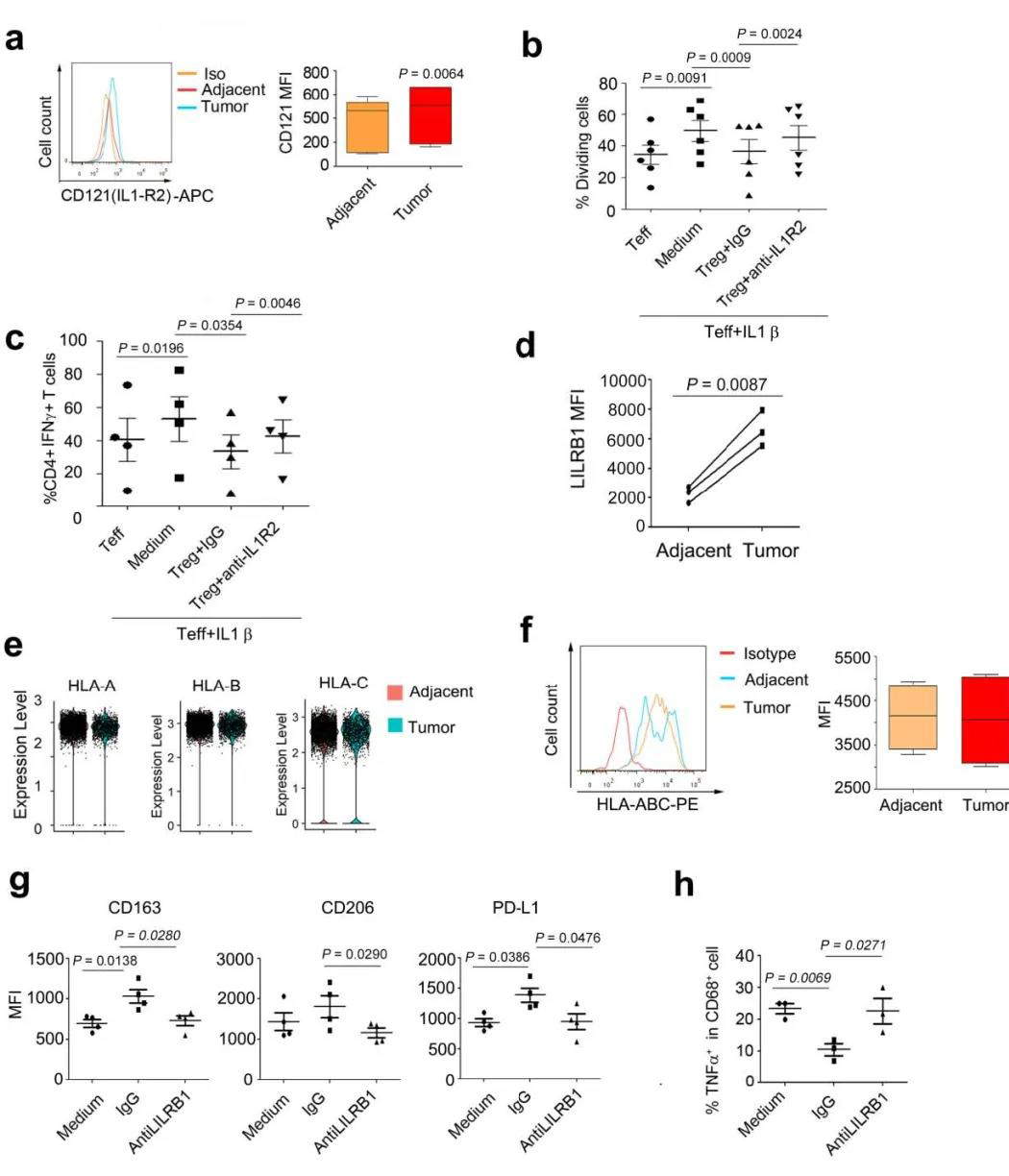
作者还预测了Tregs中的MHC和巨噬细胞中的LILRB1之间的相互作用(图7G)。MHC受体LILRB1是髓样细胞活化的负调节剂,是M2抑制状态的启动子。MHC-LILRB1相互作用抑制巨噬细胞,是癌症免疫治疗的靶。首先通过scRNA-seq分析了巨噬细胞中LILRB1的表达,并通过FACS对其进行了进一步验证。发现,与邻近组织相比,ESCC巨噬细胞中LILRB1的表达增加了(图7H-I和补充图11D)。Tregs表达高水平的HLA-A,HLA-B和HLA-C(补充图11E-F)。多色IHC染色还显示了LILRB1表达巨噬细胞和Treg之间潜在的物理相互作用(共定位)(图7J)。当Tregs与巨噬细胞共培养时,作者发现Tregs促进了表达M2标记(包括CD163和PDL1)的巨噬细胞,并降低了TNFα的表达。但是,LILRB1抗体阻滞抑制了这些影响(图7K-L和补充图11G-H)。这些数据表明,Tregs可能通过HLA和LILRB1相互作用调节巨噬细胞功能,而阻断该途径可能会促进ESCC的抗肿瘤免疫力。
总结:文章从ESCC和邻近组织获得的免疫细胞转录图谱为理解免疫状态提供了框架,并揭示了ESCC环境中免疫细胞的动态特性。此外,从许多方面说明了ESCC的免疫抑制状态,所有这些都是开发ESCC和其他癌症的免疫疗法的潜在新靶标。
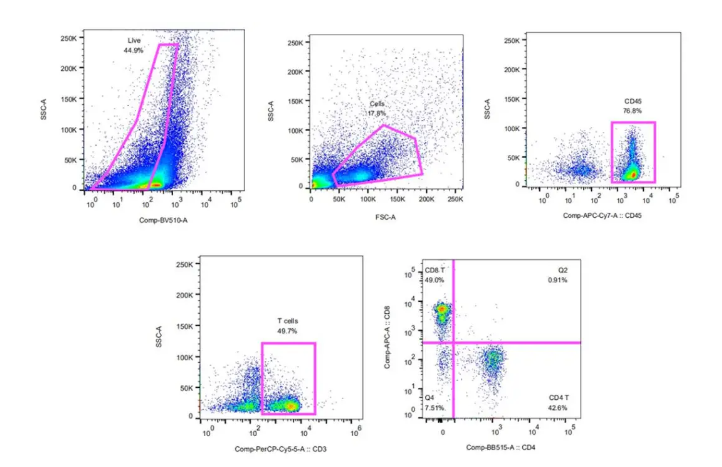
本文涉及技术: 流式细胞术
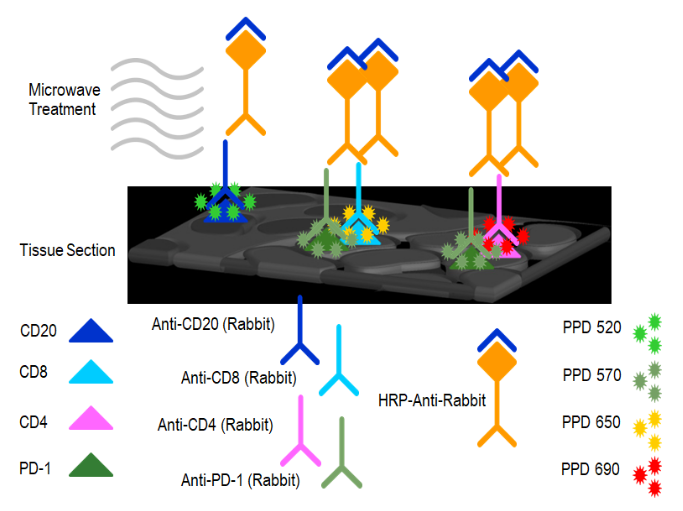
本文涉及技术: 多重免疫组化
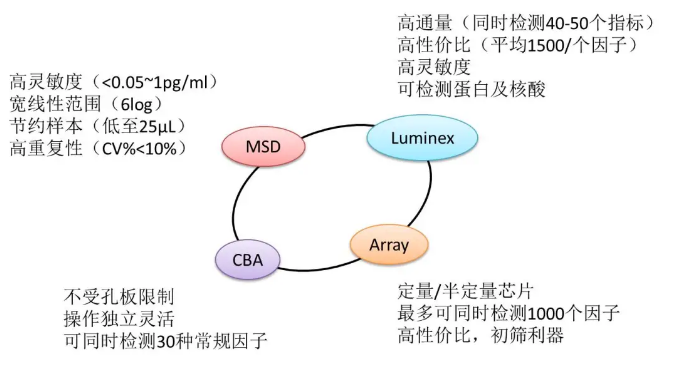
本文涉及技术: 单/多因子检测
|












 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




